Per il neurologo il martelletto è prezioso.
L’esame neurologico era considerato uno strumento diagnostico potentissimo nell’era pre-TC/RM, quando i grandi clinici del passato riuscivano a diagnosticare una lesione nel sistema nervoso spesso con stupefacente accuratezza.
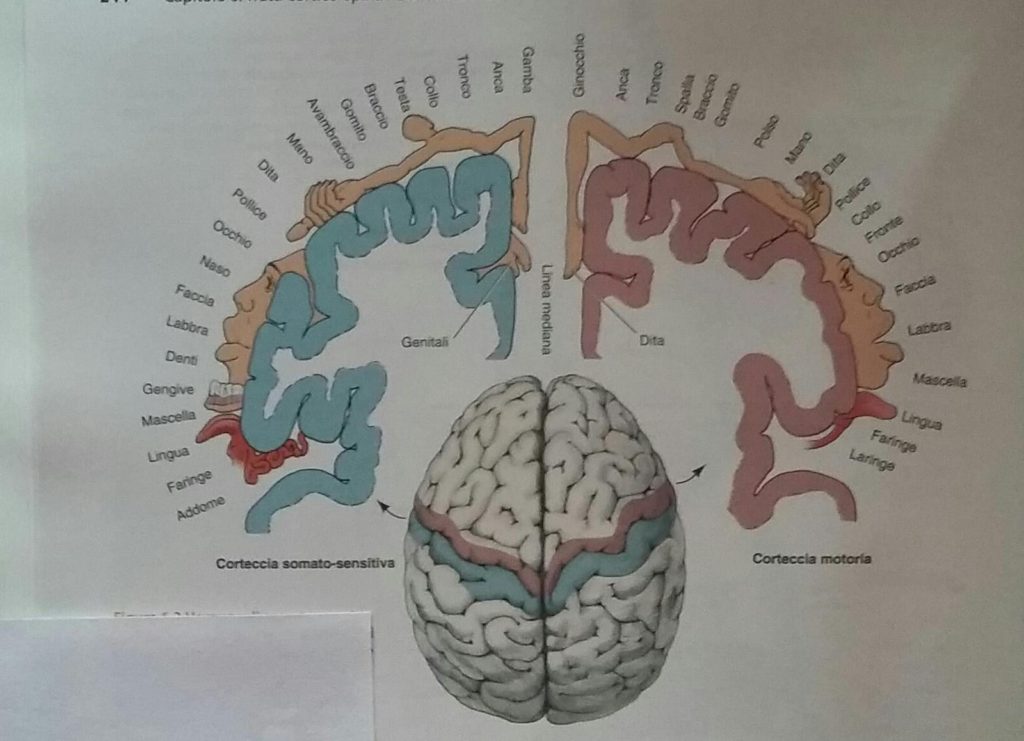
Nel corso di un esame neurologico, lo specialista, battendo il martelletto su di un tendine, evoca un riflesso tendineo profondo o riflesso mono-sinaptico da stiramento.
Questo riflesso è un arco riflesso molto noto che inizia dai recettori o fusi neuromuscolari, i quali sono capaci di trasdurre il grado e la velocità di stiramento nei muscoli in energia nervosa o potenziali d’azione. Il recettore trasduce e trasmette questa informazione ai dendriti dei neuroni sensitivi che la convogliano, lungo tutto il neurone, fino al midollo spinale. Giunti con i loro assoni nel midollo spinale i neuroni sensitivi formano sinapsi multiple, ed anche sinapsi dirette sul motoneurone o 2° neurone di moto (parte anteriore del midollo spinale, mentre dalla parte posteriore è entrata la fibra sensitiva). L’effetto di questa sinapsi diretta (neurone sensitivo, eccitato dal fuso neuromuscolare stirato, con il secondo neurone motorio) è la contrazione del muscolo.
Tra il neurone sensitivo e quello motorio ci sono anche interneuroni ed, inoltre, l’informazione sensitiva, entrata nel midollo, abbiamo visto che inizia il suo viaggio verso la corteccia cerebrale per lasciare che neuroni superiori, fino al primo neurone di moto, possano assumersi il controllo del movimento.
I nostri sistemi nervosi si sono sviluppati sullo stesso modello dei vermi, comparsi circa mezzo miliardo di anni fa. In questo inimmaginabile tempo, l’unica cosa che cambiava era il numero degli interneuroni che si frapponeva tra un numero abbastanza costante di neuroni sensitivi e motoneuroni.
Apprenderemo che l’unico vantaggio funzionale è stata l’INIBIZIONE.
Tutte le abilità che caratterizzano la nostra specie non potevano comparire se non si fosse verificata, da parte del neurone motorio centrale, un’ inibizione sulla risposta (le funzioni esecutive rappresentano questo concetto di inibizione nella sua massima espressione fisiologica). Questa funzione inibitoria, a sua volta, non poteva organizzarsi se il segnale sensoriale non avesse subito, durante il suo viaggio dalla periferia alle cortecce, un processo di modulazione e di integrazione.
Per questo motivo dobbiamo ritornare allo studio delle strutture anatomiche situate al di sopra del midollo spinale.
Al di sopra del midollo spinale troviamo il midollo allungato o bulbo che insieme al ponte ed al mesencefalo formano il tronco encefalico.
Il tronco encefalico è posizionato sotto il diencefalo (talamo, ipotalamo e altre strutture, tra cui la ghiandola pineale). Dietro ad esso c’è il cervelletto, mentre in basso, abbiamo visto, c’è il midollo spinale. Il tronco encefalico, abbiamo già visto, è importante perchè vi passano le vie che vanno verso le cortecce cerebrale e quelle che scendono verso il midollo spinale. Inoltre, abbiamo visto che è la sede dei nuclei della formazione reticolare. Dobbiamo aggiungere che è anche la sede delle dodici paia di nervi cranici. Inoltre contiene numerosi nuclei propri, che controllano i nervi cranici, il livello di coscienza, il tono muscolare, la postura e numerose funzioni essenziali quali, ad esempio, funzioni cardiache e respiratorie.
Da clinico, devo aggiungere che nessuna regione del nostro S.N.C. incute più soggezione del tronco encefalico, forse perchè piccole lesioni di questa struttura possono avere conseguenze drammatiche, o forse perchè da un punto di vista evolutivo rappresenta la più antica regione cerebrale, richiamando il cervello dei rettili.
Intorno alla linea mediana(ricordate, la parte, a mio avviso, più compromessa nei bambini con disturbo dello spettro autistico), ben al di sotto delle cortecce cerebrali, troviamo un gruppo di strutture che, evolutesi da un sistema legato all’olfatto negli animali più primitivi, espletano una serie di funzioni: la regolazione delle emozioni, la memoria, gli stimoli della fame e della sete, il controllo automatico e neuroendocrino,” il sistema limbico”.
Il sistema limbico viene avvolto dalle cortecce cerebrali che, oltre alle aree motorie e sensitive primarie precedentemente incontrate, contengono ampia quantità di corteccia associativa che può essere unimodale o eteromodale (cross-modale).
Nella corteccia associativa unimodale il processamento riguarda per lo più una singola modalità sensitiva o motoria. Questa corteccia è situata vicino all’area motoria o sensitiva primaria. Nella corteccia associativa eteromodale, invece, la funzione principale è quella di integrare modalità sensitive e/o motorie multiple (vista/udito, vista/tatto, udito/vista/tatto ecc.).
Le aree associative eteromodali, abbiamo visto, si allontanano dalle aree motorie e sensitive primarie.
Per questo motivo, abbondano anteriormente alla corteccia motoria primaria ( per questo definite anche aree motorie terziarie) e nel lobo parietale (corteccia associativa terziaria o eteromodale).
E’ interessante notare che esistono connessioni reciproche fra la corteccia primaria e associativa, nonchè tra aree motorie e sensitive (ricordate che gli strati II e III contenevano neuroni i cui prolungamenti non uscivano dalle cortecce).
Nell’ adulto, a differenza delle lesioni della corteccia primaria, le lesioni della corteccia associativa sensitiva o motoria non generano deficit severi delle funzioni motorie o sensoriali di base, ma un deficit nell’astrazione e nella pianificazione motoria.
E, in corso di neurosviluppo, cosa genereranno?